Altri klepton nel Marginella glabella Complex?
Flavio Mattavelli
Release 2011. Un sommario è stato pubblicato sul n°74 della rivista MMM (febbraio 2012). Modifiche 2013-2014-2015-2017. Ultimo aggiornamento 2021.
|
Premessa Nella scrittura sintetica talora adottata, le specie sono indicate con le lettere maiuscole come qui di seguito. Per ulteriori note sulla legenda dei simboli della scrittura sintetica, vedere anche il paragrafo simbologia sintetica. G = Marginella glabella Linneo, 1758-1767 I = M. irrorata Menke, 1828 D = M. desjardini Marche-Marchad, 1957 S = M. sebastiani Marche-Marchad & Rosso, 1979 P = M. pseudosebastiani Mattavelli, 2001 R = M. pseudodesjardini Le Béon, 2012 L = M. (kl.?) pseudoglabella Mattavelli, 2018 V = M. (kl.?) visayae Mattavelli, 2021. R è una forma morfologica identica al mio supposto "kl. incredibilis", che era in Internet nel 2011, riferitamente alla foto di un esemplare 57 mm stimato ibrido SXD. Per il futuro ho rigettato la denominazione “incredibilis”, a favore di “pseudodesjardini”, per non creare un conflitto nomenclaturale, in quanto il nome “pseudodesjardini” era già in Internet prima del 2011, sebbene solo come forma da confrontare, senza l’affermazione specifica di Roger Le Béon. Per me comunque rimane sempre il mistero dell'esistenza di possibili klepton dentro il Complex analizzato, qui solo morfologicamente. Esistono nel Complex diversi morfi "ibridi ambigui", che saranno oggetto di quest'articolo, indicando con la scrittura ...X... tutti le forme ibride morfologiche, scrittura generalmente riferibile a sia falsi ibridi, in realtà varietà subspecifiche di specie note, sia veri ibridi sterili, sia ibridi klepton ibridogenici, sia vere nuove specie per ibridazione. Ad esempio nel testo successivo “giselica” è alternativamente intesa dapprima come ibrido morfologico
PXD, poi potenzialmente rientrante nella specie R, tuttavia “giselica” potrebbe anche essere il
klepton PXR, mentre R, ibrido morfologico SXD, oltre che proprio la nuova sp.
di Roger, potrebbe anche essere un ipotetico kl.
SXD. Vi prego di scusare la possibile confusione di tutti i nomi precedenti,
che in parte verranno lasciati nella loro origine ed interpretazione storica. Per studiare meglio i morfi più grandi,
collegabili a D, S, P, R, L, tutti da confrontare soprattutto con G, nel
Complex in oggetto sono state trascurate le sp. M. goodalli, aurantia,
velai e lamarcki, che pure potrebbero essere collegate morfologicamente a
G, ma sono di dimensioni generalmente più piccole e separabili dai morfi
analizzati. Le piccole M. irrorata sono spesso facilmente confuse con G e pertanto saranno invece da considerare dettagliatamente. |
|
|
|
|
Altri klepton? Forse Marginella kl. antinea, e precisazioni riguardo alla simbologia. Biogeografia. Ulteriori klepton? "lumperia". Ibridogenesi teorico-pratica. Supposte propensioni alle ibridazioni naturali nel M. glabella Complex. Morfi RXD = DXR. Conclusione sulla rarità o frequenza delle "specie" in questione. Postilla sulle diversità tra L & G. Complex esteso a M. irrorata e cenno alle forme "albine" di G etc.; tabella fotografica riassuntiva dei morfi analizzati e loro simbologie.
|
Introduzione.
Guardando la mirabolante variabilità delle grandi marginelle (quelle lunghe oltre 30 mm) dell'Africa occidentale, l’interrogativo che si pone spontaneo è se si tratti di diverse varietà tutte della stessa superspecie (come per le innumerevoli varietà di cani), non comprendendo perché esistono tante “specie” apparentemente interfeconde, quindi un’unica superspecie, intesa come una specie molto infravariabile, che per ragioni storiche dovrebbe essere Marginella (Voluta) glabella (Linneo, 1758 e 1767?). Eppure l’interfecondità non è evidente in tutti i loro morfi, anzi alcune popolazioni si differenziano proprio come vere specie con saltuarie ibridazioni sterili. Il fatto che esistono inoltre popolazioni con ibridazioni naturali fertili è raro per il Regno animale, solitamente limitato a casi eccezionali, ma anche comuni. Ad esempio la maggioranza delle rane è ibridogenica. Anche per le marginelle del Complex di M. glabella ho tentato di introdurre il concetto di klepton, cioè di popolazione naturale ibridogenica, senza minare il concetto di specie biologica e senza trascurare la possibilità di rari ibridi naturali sterili.
Ovviamente una popolazione morfologicamente ibrida potrebbe essere una varietà di una specie nota o nuova, specie magari estremamente variabile, e non essere un klepton. Tutta la trattazione che seguirà parla di klepton, ma essi potrebbero in realtà essere sostituiti dalla voce varietà specifica, se non esistessero dubbi riguardo a quale specie far rietrare la varietà.
Purtroppo
nella maggioranza dei casi si tratta proprio di conchiglie intermedie
morfologiche, per le quali non è possibile mettere il piede in 2 scarpe, anzi
occorre costruire la scarpa adatta, presumibilmente il supposto probabile
klepton, se non probabile nuova specie.
Osserviamo che i figli somigliano ai genitori. Nulla vieta di pensare che un ibrido morfologico discenda da antenati ibridati.
Ciò non significa che un ibrido morfologico sia figlio di un’ibridazione di specie diverse, però, se c’è stata ibridazione, è probabile che il risultato somigli alle specie originarie. Dunque, quando vedo una conchiglia morfologicamente ibrida, posso pensare che discenda da quelle che riconosco come tipiche (o anche da altre specie imprecisabili, ma con minore probabilità, se riconosco nell’ibrido morfologico le caratteristiche tipiche di 2 specie note).
Ritengo che l'osservazione fatta si possa applicare ai presunti klepton, come a tutte le categorie animali, sebbene la morfologia non sia determinante per l'identificazione della categoria biologica.
Marginella (klepton?) pseudodesjardini.
Sul numero 70 della rivista MMM (febbraio 2011) ho ipotizzato la possibilità di Marginella klepton pseudodesjardini. In ogni klepton esistono 2 specie parentali ibridate, una delle quali, insieme al klepton, forma il cosidetto synklepton, genetico ed ambientale, dove il klepton può prosperare.
Non avevo precisato le specie base
del klepton perchè non è possibile certificare il klepton né il synklepton
dalla morfologia, però il supposto synklepton "pseudodesjardini" mi appariva costituito da una delle 2 specie
parentali Marginella
desjardini Marche-Marchad 1957 e Marginella pseudosebastiani Mattavelli 2001 e dal klepton di queste 2 specie fra loro
ibridate morfologicamente.
Riporto per comoditàl'esemplare
fotografato nel numero 70, senza entrare nel merito di quale delle 2 specie
parentali debba fungere da riproduttore maschio con la klepton femmina
(caso più probabile), o riproduttrice femmina con il klepton maschio (caso
geneticamente improbabile), non escludendo l'ibridogenesi tra 2 medesimi
klepton dello stesso synklepton (ibridogenesi poco probabile tranne
che per allopoliploidia, tutto da verificare geneticamente).
Se siamo in presenza di klepton, ma si tratta comunque di categorie kl. oggi non ancora ben definite per questo Complex, premetto la dicitura kl., opp. kl.? o (kl.?) nei casi dubbi, davanti al nome storico o della specie o/e della varietà, facendo slittare il nome del primo Autore tra parentesi.


Marginella (kl.?) pseudodesjardini (Le Béon) form pseudodesjardini Mattavelli 2011, oppure più probabilmente
oggi nuova specie Marginella pseudodesjardini Le Béon 2012, forma giselica Mattavelli 2013,
isola di Gorée, vicino a Dakar, centro Senegal, lunghezza 73,5 mm.
Nel 2011 sapevo che Roger Le Béon stava studiando forme molto simili, da lui denominate "pseudodesjardini", quindi io feci rientrare il morfo della foto nella stessa denominazione, supponendo in più che si potesse trattare di probabili klepton, tuttavia Roger non ha mai sostenuto l'idea dei klepton.
La categoria "pseudodesjardini", intesa in senso lato, cioè comprendente tutte le forme ibride direttamente a lei ricollegabili, pur essendo numerosa, non è numerosissima ed appare sparsa dalla penisola di Capo Verde fino al Golfo di Guinea, in simpatria con le specie con le quali appare ibridata, cioè direttamente soprattutto Marginella sebastiani, pseudosebastiani, desjardini, e forse anche M. glabella.
Nel Complex delle 4 specie Marginella glabella, sebastiani, pseudosebastiani, desjardini, resta sottinteso che, nel caso che l'idea klepton dovesse risultare geneticamente errata, esistono altre forme, meno rare di "giselica", riconducibili a "pseudodesjardini" , morfi però da intendere soprattutto di morfologia SXD, cioè i morfi tipici "ex incredibilis (vedere capitolo apposito)", quindi è possibile che esista nel medesimo Complex un'ulteriore buona specie biologica M. pseudodesjardini Le Béon 2012, come descritta nel giugno 2012 nella rivista Novapex.
Per mia convenzione mi atterrò alla scelta di Le Béon, valida fino a prova contraria.
Osservazioni ed ipotesi riguardo a "giselica".
L'esemplare di 73,5 mm era stato da me nel 2009 inteso come possibile M. pseudosebastiani form pseudodesjardini, quindi ricollegato, e forse è ancora ricollegabile, alla sp. M. pseudosebastiani, come nella seguente alternativa 1. Infatti c’è una differenza importante, riguardante la denticolatura del margine, denticolato nella tipica M. pseudodesjardini Le Béon 2012, ma nell’esemplare di 73,5 mm il margine è quasi liscio, nonostante la grande stazza della conchiglia, decisamente adulta. Pertanto occorrerebbe denominare quest’esemplare non più proprio “pseudodesjardini”, bensì ripropongo la denominazione di forma “giselica ”, introdotta in quest'articolo nella versione del gennaio 2013, oggi pienamente riconfermata come morfo distinto, per lo più ambiguamente riconducibile, se non proprio identificabile, oggi soprattutto come varietà atipica di Marginella pseudodesjardini nuova sp. di Roger, come nella seguente alternativa 2, ma lasciando aperta la strada ad altre interpretazioni, cioè precisamente la categoria di "giselica" potrebbe anche essere quella di un raro vero ibrido apparente PXD (parte dell'alternativa 3) o di un vero kl. raro, es. forse PXR (alternativa 5).
Mi sono permesso di usare una scrittura sintetica, con legenda completa della simbologia nella tabella fotografica in fondo alla pagina e precisazioni sui simboli al paragrafo "simbologia sintetica", nel capitolo inerente "...antinea = PXG = GXP".
In prima approssimazione intendere valida sempre anche la scrittura simmetrica speculare, cioè anche ad. es. PXD=DXP.
Roger
Le Béon nel 2012 ha descritto la nuova sp. R con morfologia tipica SXD,
unificando gli ibridi morfologici SXD e DXP entrambi in R.
Io nel 2011 avevo ritenute invece separate le 2 morfologie SXD e PXD, avendo chiamato allora SXD = incredibilis (oggi = R nuova sp.) e PXD = quel morfo che nel 2009 poteva essere form pseudodesjardini, ma nel 2013 fu riconosciuto diverso da R e separato in "giselica". Attualmente ritengo ancora che queste 2 morfologie vadano sempre distinte in ogni caso, sia che si tratti di kl. o di sole specie.
Nel dettaglio, per la classificazione del morfo "giselica", talora in nomenclatura trinomiale, pur incontrando alcuni problemi tassonomici, si potrebbero scrivere in totale 5 alternative:
1. Se giselica è una forma (non tipica) della specie P, si chiamerà M. pseudosebastiani (Mattavelli 2001) form giselica Mattavelli 2013, per le ragioni già esposte in pseudodesjardini.htm , ragioni che però oggi ritengo non del tutto convincenti, in quanto morfologicamente superate dalle ipotesi dell'alternativa 2 e anche della 3.
2.
Se giselica è una forma (non tipica) della specie R,
si chiamerà M. pseudodesjardini (Le Béon 2012) form giselica Mattavelli 2013, ipotesi forse la più valida in assoluto, come
indurrebbe anche il buon senso comune.
Oggi reputo "giselica" di morfologia più vicina alla nuova specie M. pseudodesjardini piuttosto che al presunto klepton PXD (pure antecedentemente
denominato "pseudodesjardini", ma con diverso significato biologico, di ibridazione di D verso
P, che oggi reputo poco probabile, a favore dell'ibridazione di D verso S),
tuttavia ho ancora dei dubbi sulla categoria biologica di "pseudodesjardini sensu R n. sp." (fatta oggi tipicamente corrispondere
esattamente all'ibridazione morfologica SXD) ed in subordine ho dei dubbi
riguardo alla tassonomia di "pseudodesjardini
form giselica". Infatti "giselica" può essere vista
soprattutto come una forma atipica di R, pur discostandosi notevolmente dalla R
tipica di Le Béon, ma, essendo quindi anche un apparente ibrido morfologico
RXP, può anche essere un raro klepton RXP (v. nel dettaglio al punto 5).
3. Occorre notare che la frequenza di "giselica" è molto bassa (nel riassunto in color ruggine a fondo pagina è indicata stimata circa 0,5%, ma di fatto l'esemplare 73,5 mm è unico), quindi potremmo trovarci anche di fronte a veri ibridi naturali saltuari occasionali sterili, tra le specie R & P oppure tra P & D. Comunque certamente "giselica", e anche gli altri morfi fertili della "pseudodesjardini sensu R n. sp.", sono poco fecondi, ma la frequenza di "giselica" è una delle più basse del Complex. Dunque, limitatamente a "giselica", la proposta di siffatta identificazione come vero ibrido non è affato da trascurare, anzi assegno (soggettivamente) all'alternativa 3 una probabilità di verità quasi massima, poco sotto a quella della 2 e poco sopra a quella del punto 1.
4. Essendo R = nuova sp., per "(kl.?) giselica" si può ancora pensare all'identificazione con PXD, ipotetico kl. di un morfo diverso dalla sp. R, morfo che potrebbe chiamarsi M. (kl.?) giselica Mattavelli 2014, se oggi denotassi una divergenza totale da R, ma (al pari di Le Béon) io non noto tanta differenza morfologica da giustificare la distinzione tra queste 2 categorie sp/kl. Questa possibilità mi appare pertanto molto scarsamente vera.
5. Le conchiglie di "giselica forse = PXD" e della sp. R=SXD sono quasi identiche, tranne che divergono nella dentellatura del margine:"giselica" ha il margine quasi liscio, mentre R è sempre dentellata dritta sul margine esterno della conchiglia di esemplari adulti. Solitamente P non è dentellata, mentre D è dentellata sinusoidale. Se sia P che R sono entrambe sp., pertanto "giselica" potrebbe essere anche (in ipotetica alternativa omonima al punto 4 = M. (kl.?) giselica Mattavelli 2014; questo nome diverrebbe definitivo solo dopo una scelta valida o del punto 4 o del 5) un altro supposto kl. PXR, per via delle morfologie comuni. Invece, sempre per le morfologie, per "giselica" escluderei un coinvolgimento diretto di S, per i seguenti motivi. Per inciso, la morfologia SXP va analizzata a parte, unitamente a quella SXG, vedere pagina Universo lumperia, e comunque il morfo "giselica" non è nè SXP nè SXG. Non ipotizzando le possibilità di esistenza del kl. SXD, essendo indirettamente R n. sp. morfologicamente già SXD, "giselica" potrebbe solo in via logica essere un ipotetico kl. SXR, ma le morfologie delle dentature presenti solitamente sia in S che in R sconsigliano anche quest'ultima ipotesi, essendo le denticolature sul labbro esterno di "giselica" assenti. Il caso 5 mi appare poco più vero e probabile del caso 4, ma sempre molto scarsamente vero e molto poco probabile, e parimenti meno probabile del caso di ibridazione n° 3.
Riassumendo, con probabilità decrescente solo basandosi su gli aspetti morfologici, abbiamo oggi per "giselica" le alternative n° 2, 3, 1, 5, 4.
Il morfo "giselica" quindi potrebbe proprio non essere un vero kl., o essere un kl. molto raro, o essere più probabilmente un ibrido casuale (caso 3). Però se si trovassero molti altri esemplari simili, anche il caso 3 perderebbe di efficacia, e potrebbe essere superato dal 5.
Morfi SXD = DXS.
Per quanto riguarda il morfo "incredibilis"
=
SXD, siccome R è ritenuta una nuova vera sp.,
certamente non si potrebbe configurare un kl., essendo il morfo identico alla
sp. R tipica. Il problema della supposta esistenza di kl. in
questo Complex, idea nata con "giselica", se
riteniamo "giselica" una
forma subspecifica della sp. R (come alternativa 2), sembrerebbe svanito nel
nulla.
Osserviamo che i morfi ibridi morfologici SXD sono molto
più comuni di "giselica"
e certamente fecondi.
Pur scartando per "giselica"
la possibilità di essere direttamente kl.,
l'idea kl. potrebbe pertanto risorgere per
i morfi SXD qualora fossero riconosciuti kl.,
annullando la qualifica di buona specie R, che è di morfologia proprio
SXD. Quindi, interpretando R come un nuovo kl.,
implicitamente verrebbe riproposta la qualifica di kl. anche
per "giselica", già
intesa secondo l'alternativa n° 2, cioè "giselica"
dovrebbe essere riproposta come varietà atipica del supposto kl. R.
"ex
incredibilis" = forma
tipica della vera M. pseudodesjardini nuova
specie.
Questo capitolo
andrebbe letto per primo, ma la storia di "giselica",
che gli è profondamente intrecciata, è cominciata prima,
morfologicamente nell'ottobre 2009, con il
morfo "giselica" individuato
allora senza il nome "giselica".
Nel febbraio 2011 (MMM n°70) "giselica" fu chiamata semplicemente ma confusamente "pseudodesjardini", allora intesa come forma innominata di un probabile kl. R. Idem
nel febbraio 2012 (MMM n°74, riassunto cartaceo della prima versione
dell'articolo Web che state leggendo).
Nel
2006 individuai un altro diverso morfo, ma allora non lo denominai affatto;
quest'altro è il morfo da me chiamato "incredibilis" nel novembre 2011,
nella versione originale di questo stesso articolo Web, dove ancora non era
citato il nome "giselica",
ma
il morfo "giselica"
era
il riferimento per introdurre il diverso morfo "incredibilis".
Ecco la necessità
di dare subito un nome, anche provvisorio e non tassonomico, alle conchiglie
che appaiono nuove.
Il fatto di questo capitolo nasce
dalla presenza di svariati esemplari, uno già citato in "pseudosebastiani.htm" e altri come
nelle foto dei siti di Le Béon (Abstract etc.e in Novapex), esemplari da lui
attribuiti dapprima a M. cfr. desjardini, poi intesa come “pseudodesjardini” nuova specie, però per me non sono “pseudodesjardini giselica”, in
quanto la loro morfologia è (invero poco) diversa.
Per me sono ibridi morfologici tra M. desjardini Marche-Marchad
1957 e M. sebastiani Marche-Marchad & Rosso 1979.
Riporto nel seguito le foto di questi che credevo rari esemplari SXD, che nel 2011 avevo chiamato forma "incredibilis", a sinistra lunghezza 57 mm, provenienza Casamance, Sud Senegal (esemplare creduto nel mio sopracitato Web un effettivo ibrido biologico occasionale M. desjardini X M. sebastiani), ed a destra un esemplare dei siti di Le Béon, desumo lunghezza circa 62 mm, morfi ascrivibili entrambi ad un'unica categoria, oggi non più rara, anche se non comune. Le puntinature chiare di “incredibilis” sono con relativamente pochi grandi punti distribuiti abbastanza caoticamente e poco allineati rispetto ai puntini tipici, in maggioranza leggermente più piccoli e numerosi, abbastanza ordinati ed allineati della “pseudodesjardini giselica”, inoltre le denticolature del margine di "incredibilis" sono evidenti, laddove in "giselica" 73,5 mm le denticolature sono praticamente assenti.
In verità nell'esemplare 62 mm le denticolature non sono evidentissime, mentre negli altri esemplari di Le Béon lo sono, ben distribuite con andamento non sinusoidale lungo tutto il margine.


Questi 2 esemplari dunque potrebbero geneticamente essere un diverso morfo della fantomatica"pseudodesjardini giselica", od anche in alternativa un altro diverso presunto klepton DXS, con synklepton basato su M. desjardini o M. sebastiani, denominabile M. (kl.?) incredibilis Mattavelli 2011 ( ipotesi tuttavia non definitiva perchè non sostenuta da prove genetiche), od infine essere proprio la nuova specie di Roger Le Béon 2012.
A prescindere dall’esemplare di 57 mm, che mi appare anche con evidente denticolatura quasi desjardinica del margine, l’esemplare di 62 mm mi appare uguale agli altri esemplari di M. pseudodesjardini di Roger, dei quali riporto un’altra sua foto nella tabella in calce alla presente mia pagina. Dunque la morfologia "incredibilis" è praticamente uguale a "pseudodesjardini Le Béon" ed "incredibilis" è quindi sinonimo di "pseudodesjardini sensu R". Oggi io, pur ritenendola valida ma non essendo ancora convinto che si tratti di una nuova specie, rilancio l’ipotesi kl. ancora senza dimostrazione genetica, però riconosco nomeclaturalmente la validità della denominazione specifica di Le Béon, come antecedente all’eventuale riconoscimento futuro dell’ipotetico mio kl. incredibilis.
Io credo di rendere in tal modo invalida la denominazione di "incredibilis" come klepton, a favore della denominazione di "pseudodesjardini" comunque intesa, onde evitare un'eventuale inutile conflitto nomenclaturale.
Se R è un klepton, incredibilis diverrebbe un sinonimo antecedente ma invalidato di Marginella kl. pseudodesjardini (ex et alias kl.incredibilis Mattavelli 2011, sp. Le Béon 2012) tramite un futuribile Autore ora sconosciuto, quello che dimostrerà la cosa in futuro, nell'anno che la dimostrerà.
Con la presente interpretazione tassonomica riconosco che già nel 2011 supposi in Internet l’esistenza del kl. incredibilis, ma non lo dimostrai, poi nel giugno 2012 Le Béon introdusse in Novapex la nuova sp. M. pseudodesjardini e, qualora venisse dimostrato in futuro che si tratti invece di un effettivo kl., il futuro Autore è pregato di aggiungere il suo nome, con l’anno della scoperta, lasciando solo la denominazione di "pseudodesjardini", senza introdurre nuove denominazioni per questo klepton.
Non so cosa ne pensi l'ICZN, ma se esistono i kl. occorrerà rivedere la sistematica linneiana definendo meglio le regole della nomenclatura.
Multiklepton.
Intendo per multikl. una ipotetica associazione di klepton oppure, se biologicamente possibile ma tutto da dimostrare, un ibrido fertile di ibridi ipotizzabili fecondi tra loro o fecondi tra sp. e kl.
I miei dubbi non riguardano solo la tassonomia formale, ma l'essenza stessa di alcune specie biologiche. Riconoscendo che P non è il kl. GXS=SXG (*), bensì una vera buona sp., ed escludendo momentaneamente la presenza legante del possibile kl.SXP=PXS, per quanto riguarda R potremmo trovarci di fronte a un multiklepton basato sui 2 ipotetici kl. (SXD) + (DXP), oppure potrebbe esistere un multikl. basato su (SXD=R sp.) + (PXR = kl.? giselica).
Se per (R sp.o kl.+ “giselica” kl.?) si tratta effettivamente di un multiklepton o di un insieme di kl. e sp., o anche sempre della stessa sp. non ibridata (ma variabile, cioè si tratterebbe sempre solo di ibridi morfologici, non ibridi reali, come precedente alternativa n°2 riguardo a "giselica", per me soluzione biologica più probabile), ai posteri l'ardua sentenza, che potranno elaborare solo dopo analisi genetiche.
Se si ammette l'ibridazione fertile tra
specie diverse, l'ipotesi klepton e forse multikl. resta
valida ed estendibile in tutto o in parte al Complex in oggetto, nonchè
forse a tutti i molluschi, in luogo della riproduzione sempre solo tra
esemplari di una stessa specie biologica.
Il problema è stabilire se, dove, come, quando, quanto numericamente e perchè scatta, sia scattata o magari scatterà l'ibridogenesi dei klepton in modo più o meno prevalente rispetto alla riproduzione tra esemplari della stessa specie, e poi soprattutto se i klepton evolvano in effettive specie future, con possibilità di riprodursi magari in multiklepton. A parte l'aspetto teorico e cervellotico delle vicende, i casi pratici per il collezionista solo morfologico restano comunque molto misteriosi.
-----
(*) Io non condivido l'opinione che, generalizzando, anche M. pseudosebastiani sia un klepton basato sulle specie parentali M. sebastiani e M. glabella, mentre potrei ipotizzare che la specie M. pseudosebastiani sia stata in passato un diverso klepton poi evoluto nella specie attuale...
Altri klepton? Forse Marginella kl. antinea, e precisazioni riguardo alla simbologia sintetica.
Nel Complex in istudio, oltre alle
specie già assodate ed al supposto dubbio klepton "pseudodesjardini giselica", ho
postulato l'esistenza di altri klepton, derivati da particolari
ibridazioni di alcune delle già assodate specie sopraddette, nel rispetto del
concetto di specie morfobiologica, principale categoria tassonomica per la
maggioranza dei molluschi, aggiungendo il concetto di klepton come categoria
secondaria, ma potenzialmente non meno importante, anzi forse origine di quasi
tutte le speciazioni dei molluschi.

 Infatti
esistono ibridi morfologici, da me in Internet ( problematica.htm
) precedentemente attribuiti a varietà sottospecifiche, esempio M. glabella antinea,
nel 2005 ibrido
morfologico ritenuto tra M. glabella x M. pseudosebastiani e
attribuito
a varietà di M. glabella, di cui
riporto pure per comodità le immagini di un esemplare lunghezza 42,5 mm (foto a
sinistra).
Infatti
esistono ibridi morfologici, da me in Internet ( problematica.htm
) precedentemente attribuiti a varietà sottospecifiche, esempio M. glabella antinea,
nel 2005 ibrido
morfologico ritenuto tra M. glabella x M. pseudosebastiani e
attribuito
a varietà di M. glabella, di cui
riporto pure per comodità le immagini di un esemplare lunghezza 42,5 mm (foto a
sinistra).
Pur non rigettando del tutto l'attribuzione ad una varietà subspecifica di M. glabella, in quanto formante una popolazione abbastanza consistente di esemplari, "antinea" potrebbe essere invece un klepton, vero ibrido ibridogenetico, forse tra M. glabella x M. pseudosebastiani, denominabile Marginella kl. antinea Mattavelli 2011, almeno provvisoriamente per parte di esemplari riconducibili in tale morfologia.
Tuttavia è anche vero che non si può
escludere anche la morfologia GXI, anzi potrebbe essere preponderante per la
maggioranza degli esemplari similari, sebbene nel 2005 e nel 2011 io non abbia
riconosciuto la morfologia GXI, ingannato dalle forme credute tutte GXP.
Procedendo nel tempo, nel 2018 e poi definitivamente
nel 2021 sono arrivato alla determinazione che esistono forme morfologiche
intermedie tra G & I, che molto probabilmente sono nuove specie sorelle di
G & I, ma potrebbero anche essere veri klepton GXI. Ho chiamato tali forme Marginella (kl.?)
visayae.
Analogo discorso deve essere avanzato
anche per un probabile klepton M. kl. mutabilis (così da me ritenuto nel 2011 l’eventuale
klepton GXI, apparente in morfi particolari, con denominazione provvisoria ed
ambigua), ma più probabilmente si tratterebbe in maggioranza di diverse forme
sempre della nuova “specie” M. (kl.?)
visayae Mattavelli, 2021, tutt’al più comprendendo, misti tra esemplari
simili, alcuni morfi di vere varietà M.
glabella mutabilis.
Precisazioni
sulla simbologia sintetica.
Si può adottare una simbologia sintetica, come legenda completa nella conclusione e come tabella fotografica in fondo alla pagina, simbologia già adottata nel n° 34 di MMM (settembre 2001) e già anticipata essenzialmente nella premessa.
Si potrebbe anche convenzionalmente scrivere per primo il componente dell’ibridazione che appare predominante morfologicamente, cioè scrivere GXP piuttosto di PXG, senza pretendere che il primo componente scritto sia anche a base del supposto synklepton, però potrebbe anche esserlo veramente, dunque la simbologia si presterebbe al riconoscimento della struttura biologica dell'esemplare, se fosse chiara la natura genetica.
Ovviamente occorrerà sempre precisare se si tratta di un effettivo synklepton, o in alternativa di un incrocio sterile di 2 specie diverse, oppure solo di una varietà subspecifica atipica (ibrida soltanto morfologica) sempre appartenente alla stessa unica specie biologica.
Ove non diversamente precisato, s’intendano provvisoriamente equivalenti le scritture simmetriche speculari, es. GXP e PXG, ed altre analoghe per altri ibridi ibridogenici o sterili o semplicemente ibridi morfologici, cioè varietà di specie non ibride, ma che appaiono come ibride.
Infatti, nel caso che tutte le ipotesi klepton risultassero geneticamente false, con questa simbologia si possono individuare comunque gli ibridi morfologici, e si potrebbe anche intendere che il primo termine è la vera specie base, mentre il secondo termine potrebbe indicare la varietà morfologica attraverso la quale la specie base appare ibridata, determinando appunto una vera varietà della medesima nota specie base, tuttavia la cosa è troppo spesso soggettiva, in prima approssimazione.
Ad esempio
per "antinea" la specie base potrebbe essere
G, ibridata morfologicamente con P, questa ritenuta soggettivamente non
biologicamente influente, quando invece potrebbe essere vero il contrario (cioè
base P), oppure anche in realtà si potrebbe trattare proprio della ipotetica
ibridogenesi GXP oppure PXG, con synklepton da stabilire.
Biogeografia.
La sp. D è distribuita in tutto il West Africa,
con prevalenza Sud Senegal, dove vivono anche i morfi R, credo in un areale però
più ristretto di quello di D e di S. La specie S si sovrappone all'areale di P,
scemando al Nord prima del confine settentrionale di P (che è Nord Mauritania),
ma proseguendo al contrario ben oltre il confine meridionale di P (che è Sud
Senegal) verso Sud. Dunque l'areale di S si estende dal Senegal compreso fino
alla Guinea e forse più a sud.
Il morfo “pseudodesjardini
giselica” (ipotizzata inizialmente PXD ma meglio
PXR o ancor meglio forma di R, localizzata circa zona Dakar = centro Senegal)
abita pertanto nella zona limite Sud dell’areale geografico di P, che è
Mauritania e Nord Senegal. Estendendo il discorso alle forme tipiche della sp.
R, queste abiterebbero anche a Sud di Dakar, dove D ed S abbondano rispetto a
G, mentre P e G scarseggiano vieppiù procedendo verso Sud.
Invece nel Nord Senegal e ancor più in Mauritania P
potrebbe legare meglio con G, colà piu comune, formando GXP, dove G abbonda
rispetto a D, alla stessa profondità ideale per P, che credo sia circa 60 ~ 90
m, profondità però forse elevata per G, e normale per D (che è stata dragata
anche a 250 ~ 300 m). Infatti “antinea” (pur diversa da G "di profondità") è quasi
assimilabile agli esemplari di G "di profondità" citati da Lipe (v.
spiegazione in inglese in "Quick reference guide",
Intermediate forms of M. glabella towards M. pseudosebastiani, inoltre
spegazione in italiano nella Guida, inoltre anche
nell'articolo "La variabilità fatta specie", allegato, Fig. 15).
Viceversa, nel Sud Senegal dovremmo trovare rare PXD e
meno rare (ma sempre poco comuni) R= DXS, pure a discreta profondità, però non
ho informazioni per affermare le profondità con certezza, né per accertare la
struttura biologica dei supposti synklepton morfologici e/o specie citate.
Infatti nel Sud del Senegal la morfologia "giselica" sparisce, mentre compare abbondante già nel Sud Senegal e più a
sud la morfospecie DXS tipicamente identificabile con la sp. R.
Più che continuare a parlare di "giselica" (perchè una rondine non fa primavera), ritengo più significativo porre l'attenzione sulla popolazione dei campioni DXS intesa come sp. R tipica, ma potrebbe essere anche kl. R tipico. La mia impressione è che l'areale di R in prevalenza sia sovrapposto a quello di S solo a sud dell'areale di P, soprattutto nella zona senegalese simpatrica dell'areale di S e D, e continui a sud nella Guinea e forse più a sud.
In contrapposizione l’areale di distribuzione di “antinea” mi appare localizzato e sovrapposto solo nella zona nord dell’areale di P, cioè le due categorie “antinea e pseudodesjardini sensu Le Béon” credo che siano a distribuzione geografica alternata (nord e sud), presenti solo ai confini rispetto ai limiti dell’areale P, inoltre entrambe mi appaiono di consistenza limitata rispetto alla consistenza delle popolazioni di P, S, G, D.
Mentre a nord potrebbe prevalere l'ibridogenesi PXG = "antinea" , a sud dell'areale di P, a mio parere prevale la specie S, nell'ipotetica ibridogenesi virtuale con D, oppure compare in realtà un'ibridazione solo morfologica, nello status della buona specie R.
Fondamentalmente occorrerebbe proprio stabilire se S è ibridata con D (ipotizzando R = kl. SXD) oppure se si tratta in verità di una nuova sp. R (sensu Le Béon, 2012) = Marginella pseudodesjardini. Purtroppo geograficamente sono possibili entrambe le soluzioni.
Un analogo discorso riguarda la nuova "specie" Marginella (kl.?) pseudoglabella = L, per la quale rimando ai miei articoli appositi.
Ulteriori klepton? "lumperia".
Alcuni ulteriori esemplari appaiono
morfologicamente intermedi tra M.
sebastiani e M. pseudosebastiani, da far
pensare che forse possa sussistere nel Complex in oggetto unquarto klepton SXP o PXS e relativo synklepton, qualora non si pensi che si tratti della
medesima specie biologica (S=P) in diverse varietà subspecifiche, oppure
qualora non si riesca ad inquadrare gli esemplari esaminati in una
varietà delle 2 specie ritenute valide.
Invero nel 2001 credevo solo ad un'ibridazione morfologica tra S,P ed eventualmente G, tant'é che nella fig. 8 di pag. 7 di MMM n° 34 avevo indicato un esemplare morfologicamente PXS, lunghezza 39,6 mm, "tribrido" di G, esemplare di incerta classificazione pensato come varietà di una delle 3 specie. Oggi credo che tale esemplare potrebbe essere in alternativa un giovanile di P, senza riconoscerlo come possibile klepton.
Successivamente al 2001, quando pubblicai in Internet la
Guida di riconoscimento rapido, indicai incerti esemplari come giovanili di P, ad esempio vedere nella Guida le foto dell'esemplare lunghezza 43 mm. Lo
scoglio dei giovanili di P confondibili con G può essere un problema, in quanto
io non conosco P lunghe meno di circa 30 mm, che pure devono esistere.
Addirittura sotto questa misura, comune alle G, si potrebbe sconfinare molto
confusamente anche nella morfologia di M. irrorata, pur senza arrivare a pensare
esageratamente che le M. irrorata potrebbero essere i giovanili di P.Vero è che l'aspetto
conchigliare cambia con l'età dei molluschi, ma le differenze tra M. irrorata e pseudosebastiani sono troppo notevoli. Le spire apicali
e soprattutto le flammule subsuturali sono diverse, i puntini di M. irrorata sono più piccoli, fitti e reticolati
(talora a zic-zac in M. irrorata, mai
in P) ed il profilo conchigliare è più tozzo di quello di P. Invece,
analogamente ai casi GXP, le differenze morfologiche tra alcune G e M. irrorata possono essere minime. Allora è
ipotizzabile un'ibridogenesi anche tra G & irrorata? Vedere la tabella di foto a fine
pagina, che sembrerebbe confermare questa supposizione, ma per ora lasciamo M. irrorata fuori discussione. Limitiamoci ad
analizzare marginelle di dimensioni mediamente superiori a circa 30 mm.
Ecco qui sotto la foto di un esemplare,
in passato dichiarato M.
sebastiani atipica,
con i punti piccoli, probabilmente anche giovanile
di S, che ora potrebbe anche essere interpretato
invece come un virtuale ibrido ibridogenetico SXP o PXS.
|
|
In
realtà la distinzione tra le specie S & P è in generale evidente,
quindi i casi particolari di ibridazione morfologica SXP sono da attribuirsi
o a pochi esemplari atipici di una delle 2 singole specie, magari
giovanili, o a veri ibridi per lo più sterili, o forse ad effettivi ibridi
ibridogenetici, cioè klepton, se venissero riscontrati esemplari in numero
rilevante, cosa che al momento non è. Eventuali veri SXP sono quindi rari e
con troppe caratteristiche individuali, da non poter inquadrare
un'unica categoria significativa. Nel
limbo degli esemplari di incerta classificazione morfologica dunque mettiamo,
oltre ai giovanili di P che possono venir confusi con G ed oltre alle atipiche
forme di S che possono venir confuse con P, mettiamo i virtuali PXS = SXP. Se volessimo dare un nome a questi PXS etc., propongo "Marginella lumperia 2013" come "nomen dubium", come un limbo unificante varie tendenze ibride non ben identificabili in un'unica popolazione di morfi analoghi e diversamente ambigui. Potremmo includere nello stesso limbo anche altre 2 combinazioni logiche forse pochissimo realizzate in natura e qui non raffigurate, cioè GXD = DXG e GXR = RXG. |

Per
approfondire tutti questi casi ibridi ambigui, soprattutto PXS, ma poi
prepotentemente SXG, ho sviluppato la pagina Universo
lumperia. Infatti per quanto riguarda l'ibrido morfologico GXS = SXG
(ibrido che beninteso non è P!) è poi emersa la conoscenza
di L (che beninteso non è G!)
Ibridogenesi teorico-pratica.
Nel caso delle rane la consistenza di una popolazione klepton è numericamente paragonabile al numero di presenze della specie base del synklepton, anzi talora i klepton sono addirittura in numero superiore, in un vasto areale, mentre in queste marginelle, posto che esistano klepton misti a vere specie, a prescindere dai ridotti ed incerti casi di cui sopra, anche negli evidenti casi di “pseudodesjardini & antinea”, i supposti klepton appaiono localizzati geograficamente e poco numerosi rispetto alle popolazioni delle specie base dei supposti loro synklepton. A parte il riconoscimento di un’ibridogenesi limitata, resta da stabilire nei singoli casi se si tratta di klepton in fase evolutiva iniziale, o stabilizzata nel tempo (klepton permanenti?) o in fase quasi speciata in una nuova specie, escludendo che si tratti di varietà specifiche.
Probabilmente
le rane attuali sono a ibridogenesi stabilizzata, mentre “pseudodesjardini
& antinea” sono solo in fase iniziale…
Rammento
che la questione dell'esistenza dei molluschi klepton non può essere
risolta su base solo conchigliare.
Tuttavia, in mancanza di informazioni
genetiche, tenuto conto degli areali di distribuzione e soprattutto delle
frequenze numeriche delle varie categorie di esemplari, a prescindere
dalle forme fossili, avendo effettuato osservazioni su un vasto numero di
conchiglie del complex di M. glabella, la distinzione tra le forme
tipiche di M. glabella, desjardini,
sebastiani, pseudosebastiani è
qualitativamente ma anche quantitativamente evidente, quindi non si tratta
sempre della stessa unica specie, o meglio categoria, morfobiologica, ma
di un complex di 4 ben notevoli popolazioni ognuna ben distinguibile e
pertanto inquadrabili in 4 distinte specie
parentali, tuttavia simili, geneticamente vicine e facilmente
ibridabili in particolari condizioni biogeografiche,
potendo in qualche caso determinare popolazioni numericamente inferiori ed
inquadrabili nella categoria biologica klepton, a giudicare dal consistente
numero (tuttavia ben inferiore al numero degli esemplari delle 4 popolazioni
specifiche principali) degli esemplari fra loro simili e con caratteristiche morfologiche
intermedie a quelle delle 4 specie, senza poter ulteriormente distinguere morfi
ibridi degli ibridi, né poter attribuire le consistenti popolazioni inferiori a
varietà delle 4 specie note (perché sarebbero varietà troppo
morfologicamente ibridate da non poter attribuire un’appartenenza specifica
univoca indiscutibile).
Ragionando al limite, le popolazioni inferiori inquadrabili come varietà discutibili delle 4 specie note non sarebbero inquadrabili come rare specie nuove, avendo in parte caratteristiche morfologiche delle sp. già note e, qualora non avessero nuove caratteristiche proprie, a fine discussione, se non fossero riconosciute varietà delle sp. note, potrebbero e, se lo fossero, dovrebbero essere tutte riconosciute essere solo klepton, o multikl., avendo scartata l'esistenza di un'unica superspecie suddivisa in sp. e sottospecie, con possibilità di tante introgressioni genetiche comuni tra i componenti dell'ipotetica superspecie.
Nel caso di "pseudodesjardini sensu Le Béon = ex incredibilis" non mi appaiono caratteristiche diverse da quelle di S, D e P, quindi il morfo potrebbe essere un kl., ma la morfologia è talvolta ingannevole, dunque riconosco che R possa essere una nuova sp. "sorella", anzi è così per mia convenzione provvisoria.
Nella maggioranza dei casi a
morfologia ibrida, quando si tratta di pochi esemplari fra loro simili, io
credo che siano ibridi sterili, mentre quando si tratta di esemplari numerosi,
ad es. nei discutibili casi delle "pseudodesjardini sensu Le Béon = ex incredibilis" ed “antinea”, io credo comunque che possano
essere intesi anche come categorie biologiche presunte ibridogeniche, da
verificare geneticamente, per escludere che si tratti di specie nuove, o
varietà di qualche specie già nota.
Ovviamente
potrebbe confermarsi il dubbio che anche le cosidette specie note siano in
realtà tutte o in parte derivate da klepton o essere esse stesse klepton in
evoluzione.
Non so se si possono
perpetuare gli ibridi dei (supposti) klepton, magari con
fusione in un futuro unico "superklepton", oppure se si amplierà
la biodiversità con future speciazioni geneticamente distinte, ma credo sempre
più morfologicamente indistinguibili, come nei casi delle altre "sister
species" del Regno animale.
Ad ogni modo,
senza intenzioni "splitter", credo positivo aver praticamente
distinto tutte le diverse morfologie, anche per evitare che si confondano i
morfi ibridi degli ibridi in un calderone "lumper" che non gioverebbe
a chiunque voglia scoprire la verità del Complex in oggetto.
Supposte propensioni alle ibridazioni naturali nel M. glabella Complex.
Le considerazioni seguenti non contemplavano la presenza della settima "specie" M. pseudoglabella = L (che prima del 2017 poteva essere nel limbo SXG). Vi prego in realtà di considerarne l'esistenza, anche se ho lasciato immutato il testo seguente, per diminuire le variabili in gioco (artificio non verosimile in natura).
<< Constatazione sperimentale.
Le 6 combinazioni teoriche di possibile ibridazione morfologica delle 4 specie suddette (DXP=giselica, DXS=incredibilis, PXG=antinea, PXS=lumperia, DXG e SXG) non sono tutte fortemente realizzate in natura, a parte PXG e DXS=R, che invece è talmente notevole e abbastanza abbondante da dedurre che sia proprio una vera quinta buona specie, pur essendolo già per mia convenzione provvisoria.
Sintesi pratica supponendo possibile l'esistenza di klepton tra le sp. G, S, P, D.
La situazione naturale offre in realtà fecondi solo i 2 presunti kl. e numerosi ibridi morfologici SXD (pseudodesjardini sensu R= incredibilis, poi ritenuto sp. ed ora per assurdo kl.) e GXP (antinea, prima ritenuta varietà di sp. e poi presunto kl.), a parte il dubbio morfo PXD o PXR che sia (pseudodesjardini giselica, inteso possibile kl. raro ma soprattutto morfo coinvolgibile come forma rispetto alla sp. R, questa però qui intesa come possibile kl., magari anche in un ipotizzabile multikl., al momento indefinibile), mentre le altre 3 combinazioni teoriche, PXS, GXD, GXS, (+ GXR?), sono solo sporadiche, con minori presenze e con discutibili esemplari ibridi di incerta ed opinabile identificazione morfologica ("lumperia"), quindi, per i modesti pesi di presenze, le potremmo considerare fuori gioco.
Ho leggermente ridimensionato quel che
avevo scritto nel 2001 sulla rivista MMM a proposito della frequenza degli
ibridi, ed in questo stesso articolo nel 2011 e nel n° 74 di MMM del febbraio
2012 (le frequenze di SXD e PXD erano errate e ora sono state corrette), avendo
ora constatato una maggior presenza di “pseudodesjardini
sensu R”, con piccole differenze di frequenza degli
altri “ibridi”, differenze per lo più dovute alla loro rarità, che confonde
calcoli precisi.
La sp. P appare attratta dalle specie fra loro più lontane, G & D, mentre, per la scarsità di esemplari, sembra che esista una repulsione tra P e la specie a lei più simile, S.
Invece G& D presentano un
comportamento normale per specie lontane, nel senso che si respingono fra
loro, ma possono legare con specie vicine, in particolare con
P, tuttavia G è meno legata con S (a meno che non s'intenda GXS = P,
intendimento che potrebbe essere inteso morfologicamente esatto, ma per me è
errato biologicamente, fino a prova genetica contraria), mentre D appare molto legata con S, intendendo SXD = pseudodesjardini sensu R, intendimento
che per me è morfologicamente esatto, ma biologicamente è molto dubbio. R potrebbe essere
sia una nuova sp. che un kl., klepton soltanto in questo capitolo elaborato per assurdo.
Forse
P può essere ambientalmente più ibridabile delle altre specie,
comunque è un'illazione arbitraria personale, non suffragata da dati genetici. L'attribuzione della specie parentale
P a base dei synklepton PXD e GXP è totalmente arbitraria, derivante dalla
mia impressione che P possa essere ambientalmente più ibridabile
delle altre specie, e che, ove manchi P, a Sud di Capo Verde, possa subentrare
S negli eventuali incroci SXD e GXS, incroci che oggi ho
rilevato essere più frequenti di quanto scritto in precedenza.
Ciononostante S appare la specie meno ibridabile nel complex P, G, D, S (senza aggiungere al complex anche Marginella goodalli, specie con la quale talora S sembra ibridata), soprattutto considerando che l'ibridazione morfologica SXD potrebbe essere in realtà proprio la specie R, quindi in quanto tale geneticamente indipendente, ma teoricamente ibridabile verso le altre 4 specie, ed in questo caso di R nuova sp., S consegue essere poco ibridabile. Considerare però che la sp. S è molto variabile (v. Guida riferimento).
In effetti la ora posta vera sp. R mi appare ibridabile verso P (v. giselica), verso D (v. sotto) e verso S (da verificare se esistono forme "mixed & elongate" di S direttamente collegabili a R), e anche verso G (v. Universo lumperia), però sono tutti casi discutibili e rari, quindi assegno alla sp. R un'ibridabilità specifica apparentemente alta, ma relativamente scarsa. Forse non si tratta di vere ibridazioni, ma solo di forme di ibridazione solo morfologica, in realtà subspecifiche di R (o della sp. collegabile), o ibridi occasionali. In ogni caso la variabilità di R non mi appare elevata.
Invece nel caso che R fosse un kl., il discorso potrebbe cambiare radicalmente, sia per R che per P.
S dovrebbe venir considerata essere molto ibridabile, soprattutto nei morfi SXD (ma potrebbe anche ibridare proprio notevolmente in SXG).
R sarebbe il risultato dell'ibridazione ibridogenetica tra S e D.
Anche il risultato dell'ibridazione morfologica tra S e G potrebbe venir sconvolto, quest'ipotesi potrebbe non essere più considerata rara, qualora venisse riscontrato che SXG fosse un kl. originante P, che decadrebbe dalla qualifica di nuova sp., divenendo un futuribile nuovo kl.
Attualmente comunque credo provvisoriamente che P & R siano entrambe effettive vere buone specie biologiche, restringendo il concetto di klepton "in fieri" a tutti gli altri morfi "ibridi ambigui" analizzati nell'articolo, ovviamente escludendo, ove possibile, i casi degli effettivi veri ibridi naturali occasionali saltuari sterili.>>
Post Scriptum (aggiornamento 2017): Un discorso analogo può valere per M. pseudoglabella, intendibile come klepton G X S, ma attualmente comunque credo provvisoriamente che L possa anche essere un'effettiva vera buona specie biologica, dato l'elevato numero di esemplari (circa una trentina) riscontrati come sintipi nel sito Web di Conchology Inc. ed anche oltre un'altra trentina di varietà di L a me note, senza contare quelle riscontrabili in futuro in altre collezioni, da rivedere con occhio scaltrito e attento. Tuttavia oggi L sarebbe una "specie" ancora abbastanza rara, se non gli fosse riconosciuta l'appartenenza di alcune denominate erroneamente G bordeaux, quindi l'argomento è discutibile e purtroppo difficilmente risolvibile solo morfologicamente. Vedere l'articolo Nuovo sguardo ai mondi di M. glabella e pseudoglabella.
Morfi RXD = DXR.
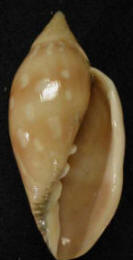
 Propongo
le immagini recto e verso, "dealer" Cochlodesma, di un raro (ma non
unico) ibrido DXR, lunghezza 73 mm, località Popenguine (Senegal), morfo adulto
che potremmo chiamare "eurekia", presentante punti chiari
anche nelle bande chiare, e non presentante la dentatura sinuisoidale, 2
caratteristiche che lo distinguono dalla morfologia tipica di D. Invece il
morfo è collegabile a R anche se il labbro esterno è liscio, ma con un'impronta
centrale, forse per via di un trauma occorsogli in gioventù.
Propongo
le immagini recto e verso, "dealer" Cochlodesma, di un raro (ma non
unico) ibrido DXR, lunghezza 73 mm, località Popenguine (Senegal), morfo adulto
che potremmo chiamare "eurekia", presentante punti chiari
anche nelle bande chiare, e non presentante la dentatura sinuisoidale, 2
caratteristiche che lo distinguono dalla morfologia tipica di D. Invece il
morfo è collegabile a R anche se il labbro esterno è liscio, ma con un'impronta
centrale, forse per via di un trauma occorsogli in gioventù.
Per questo ibrido morfologico DXR si potrebbero avanzare 5 alternative analoghe
a quelle già avanzate per "giselica",
conducenti alla fine a quasi analoghe deduzioni di identificazione.
Per lo più basta sostituire P con D nelle alternative, ottenendo alla fine una
successione simile di probabilità decrescenti (analoghe ai n° 2, 3, 1, 5, 4),
cioè soprattutto "eurekia"
potrebbe essere una forma non tipica ma
subspecifica della sp. R , poi con minor probabilità un
ibrido effettivo, poi una forma di D non tipica, poi molto difficilmente un kl.
DXR e poi falsamente un altro fantasioso kl. tra D e S
oppure tra D e P, falsamente perchè gli ultimi 2 casi sono già stati
identificati rispettivamente essere la sp. R oppure essere "giselica". Da un
confronto diretto delle conchiglie "eurekia
e giselica" infatti divergono per la forma della cuspide e per
la puntinatura dell'ultimo giro. Ovviamente se invece R fosse un kl., "eurekia"
potrebbe essere un'altra varietà diversa da quella del supposto kl.
tipico DXS.
Conclusione sulla rarità o frequenza delle "specie" in questione.
Ricordo che talora indico con "specie" una categoria biologica della quale non posso sapere morfologicamente se si tratta di vera specie oppure klepton, escludendo che si tratti di veri ibridi, ma categoria che ciò non ostante riesco morfologicamente a distinguere e separare, pur non ritenendomi uno "splitter".
Credo che M.irrorata abiti negli stessi luoghi e con la stessa frequenza di G, mentre M. goodalli genericamente in Senegal e forse con la frequenza di P, ma non ho trovato dati certi, quindi ho provvisoriamente escluso M. irrorata, goodalli ed altre piccole ritenute effettive specie fuori discussione dal successivo elenco riepilogativo dei più grandi morfi più simili per ibridazioni morfologiche, elenco che però potrebbe essere ulteriormente limitato al principale gruppo (tetraedro morfologico di vertici) G, S, P, L, comunque per necessità analizzando a parte anche il gruppo D & R e suoi ibridi col suddetto gruppo principale tetraedrico. Anche M. irrorata, goodalli, etc. potrebbero "kleptonizzare", ma ho escluso dette specie anche soprattutto per semplificare l'elenco riepilogativo.
Gli eventuali ibridi tra le specie di un gruppo potrebbero sussistere o all'interno del gruppo o verso gruppi di specie esterni, rubando percentuali di presenze ai valori dei vertici delle "specie" analizzate. Aumentando il numero di vertici, le singole percentuali di frequenza devono diminuire, lasciando il totale 100%. Ecco allora una simulazione, solo per la parte di Complex che presenta i morfi più grandi, morfologicamente vicini e confondibili.
L'introduzione di M. pseudoglabella sconvolge ogni precedente elenco di frequenze, aprendo questioni forse irrisolvibili riguardo alle frequenze dei suoi possibili ibridi, in funzione della impossibilità morfologica del loro riconoscimento certo.
Ho considerato in tot. 19 morfi caratteristici, raggruppando le loro varietà ed elencando la percentuale di frequenza delle singole categorie, attribuendo loro un valore percentuale virtuale derivato da mia impressione, sul totale 100% di presenze in tutto l'Atlantico West Africano.
Le specie basi dei possibili synklepton
restano sempre comunque indefinibili. Oggi, dopo il lavoro di Roger, anche il
supposto kl. SXD è ritenibile, in alternativa più accettabile ma da dimostrare,
una vera nuova sp. R. Ho semplicemente invertito le frequenze di PXD e SXD
scritte nel 2011 e nel n°
74 di MMM del febbraio 2012, in adeguamento all'introduzione di R sensu
Le Béon, tuttavia
le reali percentuali di frequenze potrebbero essere ancora diverse.
Ripeto che si tratta di stime numeriche puramente orientative riferite a tutto il West Africa, ma occorre tenere conto delle distribuzioni geografiche locali, dove le singole "specie" sono effettivamente presenti, quindi le frequenze vanno intese concentrate nelle zone indicate.
.
Da notare che GXS e PXS sono praticamente indistinguibili, sebbene GXS sia caratterizzata da un maggior colorito di fondo e da una puntinatura chiara leggermente meno distinta di quella di PXS. Nel 2014 ho attribuito a questi 2 morfi una frequenza totale quasi 4%, valore molto arbitrario. Se poi si considera GXS = L nuova "specie" Marginella pseudoglabella Mattavelli, 2017, detta frequenza è invero bassa rispetto alle frequenze medie delle altre ritenute vere specie. Considerando che una parte delle conchiglie in precedenza ritenute erroneamente G bordeaux (o "dark G") dovrebbe invero confluire scorporata nella nuova L, immaginiamo spostando circa un 4% del totale, ed aggiungendo circa un ulteriore 2% per la popolazione senegalese dei sintipi di L 2017, quindi spostando circa in tot. 6%, la percentuale della frequenza di presenza di tutte le varietà di L tipiche, atipiche e dubbie PXS salirebbe al 10% del Complex analizzato, mentre la percentuale delle vere G scenderebbe al 18%. Gli esemplari di L sono ancora relativamente pochi per trarre conclusioni certe, ma la loro separazione dalle altre specie è innegabile.
Gli ibridi di L saranno stimati aleatoriamente, come "lumperia imprecisabili sterili, o possibili eventuali klepton o varietà di specie fertili", sottraendo le loro percentuali da quelle del morfo stimato più vicino, preesistente nel passato elenco, che non contemplava morfi di L ibridati. Gli ibridi di L sono teoricamente possibili, ma non ancora praticamente riscontrati con certezza: potrebbe trattarsi solo di varietà atipiche delle "specie" coinvolte, magari in realtà solo con ibridazione apparente.
In tutti i casi s'intenda la reciprocità AXB = BXA degli ibridi, contemplati tutti come ibridi morfologici nell'elenco che segue.
.
G = Marginella glabella, specie comune dal Marocco al Senegal, molto diversificata in molteplici varietà di popolazioni (dark?, nane, mutabilis, albida, ghepardica etc.), conteggiando tutte le varietà eccetto i morfi seguenti, frequenza totale 24% - 6% ipotetico senza parte delle dark ed allo scopo di considerare la presenza delle L 2017, tot. G effettive = 18%
GXP = probabile kl. antinea,
Mauritania e Senegal, poco comune, morfi diversificabili in tipico
“antinea” e atipico “problematica” ( problematica.htm ), frequenza totale
era in passato 5% - 2% oggi attribuito a LXP, frequenza residua di GXP = 3%
LXP = in prima approssimazione 2%
LXG = per semplificare frequenza posta 0%, pensando di distribuire apparenti ibridi in realtà o solo varietà di L oppure solo varietà delle G scorporate da L.
L = GXS = "pseudoglabella, 2014", ibrido morfologico biologicamente da classificare, poteva arrivare a frequenza 2% + 6% ipotetico per ex parte delle dark G e per la popolazione effettiva dei sintipi di L 2017. Nel 2017 questo morfo è stato da me ritenuto nuova "specie" perlopiù del Senegal: vedere qui, quindi la presenza tot. delle L effettive in tutto il West Africa (senza forme PXS) = 8%
LXS = in prima approssimazione 1% (percentuale sottratta al passato valore di S).
GXD, GXR, LXD e LXR sono ibridi morfologici stimo difficilmente ibridogenetici, più probabilmente possono essere veri ibridi sterili, o rare forme individuali non riconoscibili come varietà né synklepton, Mauritania e Senegal, aggiungendo i veri ibridi sterili di questo Complex, frequenza totale circa 1%.
P = Marginella pseudosebastiani,
specie comune in Mauritania e meno in Senegal, poco diversificata in varietà
atipiche, frequenza 16%.
PXD = "pseudodesjardini form giselica", Sud Senegal, raro morfo tipico con puntini piccoli e allineati, frequenza circa 0,5%, oppure
RXP = altra interpretazione dubbia di "giselica", interpretazione che potrebbe vanificare la precedente, lasciandone immutata la sostanza.
RXS = teoricamente possibile, ma non ho trovato esemplari decisivi con tale morfologia.
RXD = "pseudodesjardini form eurekia", Senegal, raro morfo con puntini anche nelle bande chiare, frequenza circa 0,5%.
PXS = parte di "lumperia, 2014" , morfi virtuali da classificare, confondibili con esemplari giovanili di P ed L atipiche, era stimata 2%. Senza contare il nucleo dei morfi già caratterizzato in "pseudoglabella GXS insieme a tutte le vere L 2017", la frequenza parziale delle restanti "lumperia imprecisabili fertili" potrebbe restare al 2%, perché eventuali klepton di L con P o S sono stati stimati, assieme a possibili veri ibridi, nelle voci LXP oppure LXS.
SXD = R = "pseudodesjardini"(ex “incredibilis”),
ibrido morfologico oggi specie M. pseudodesjardini Le Béon, 2012, Sud
Senegal, frequenza 7%.
S = Marginella sebastiani, specie comune in Senegal e Guinea, molto diversificata (forme lellia, elongated etc.), frequenza totale 22% - 1% = 21%, se trovassimo veri ibridi o klepton SXL.
D = Marginella desjardini, specie
comune dalla Mauritania alla Costa d'Avorio, con notevoli variazioni
individuali, tuttavia senza dare varietà configurabili in popolazioni
nettamente diversificabili dal morfo D tipico (cioè tutte le D sono facilmente
identificabili come tali, pur presentando notevoli variazioni morfologiche
individuali), frequenza 20%.
.
I veri ibridi sterili potrebbero avere al massimo la somma delle percentuali di GXD, GXR, LXD, LXR, PXD e RXD, cioè arrivare ad una frequenza totale 2%, più una parte del 3% assegnato in prima approssimazione agli altri 3 eventuali ipotetici ibridi morfologici di L (quindi tot. max. 5% se non esistessero casi klepton o varietà di specie confuse per ibridi, tuttavia io credo che proprio varietà di "specie" fertili potrebbero essere la maggioranza di tale 3%, nei 3 casi LXP, LXG, LXS).
Infatti non ritengo sterili le "antinea" = GXP e le "pseudoglabella" = GXS (frequenza 8%) assieme ad alcune "lumperia" = PXS (fertili stimate 2% del totale delle frequenze).
La somma delle frequenze di GXS + PXS offriva nel 2014 una frequenza totale 4%, elevata nel 2017 al 10% per contare la presenza di L etc. come sopra precisato.
Le affinità morfologiche della quaterna fertile G,P,S,L mi inducono a credere nella fertilità di LXP, LXG, LXS, ma la cosa è dubbia. Vedere successiva Postilla.
Sul totale del Complex analizzato restano quindi non ben classificabili i veri ibridi occasionali sterili, di fatto tra il 2% e credo molto meno del 5% (vedere un criterio di separazione al capitolo Applicazione in Specie/klepton, dove e quando tuttavia L non era ancora considerata, quindi non considerare il suddetto 3% di differenza, o meglio di imprecisione di attribuzione di alcuni esemplari ibridi morfologici di L).
Anche purtroppo in base ai limiti di capacità di individuazione da parte dell'osservatore puramente morfologico, le 2 categorie biologiche di importanza certa e più numerose di esemplari, cioè le vere sp. oppure i veri kl., 2 categorie pur morfologicamente distinguibili e distinte unitariamente da ibridi morfologici dubbi, restano comunque biologicamente non qualificabili nella loro essenza di "specie" biologica, unitariamente distinta dalle altre "specie" del Complex.
La variabilità della specie D richiama l'elevata variabilità di G e di S, mentre P ed R sono poco variabili, ma mi appaiono assai ibridabili, anzi probabilmente sp. ibridogeniche, possibili componenti di kl., come G. Forse addirittura P & R sono esse stesse discutibili categorie klepton.
La specie G appare legare molto, ma solo morfologicamente, anche con M. irrorata, sebbene tra queste 2 specie sia già stata assodata nella letteratura scientifica una netta differenziazione biologica, risultante dallo studio delle parti molli.
Invece G appare inestricabilmente legata a L, forse come 2 "specie sorelle", se L non è un "klepton (GXS?) fratello" degli altri ipotizzabili e discutibili klepton nel tetraedro morfologico G, S, P, L. Non lo sapremo mai senza analisi genetiche di tutti i molluschi.
Postilla sulle diversità tra L & G (e ibridi collaterali).
Le possibilità di confusione morfologica
tra le vere L e le vere G sono tante
e difficilmente risolvibili,
considerando, oltre alla elevata variabilità di tutte le vere G, la pur elevata
variabilità di L. Ciò potrebbe far supporre che G & L siano altamente
ibridabili, oltre che tra sé stesse, anche con quasi tutte le altre specie
vicine dello stesso Complex, ma il fatto che sussista un’ibridazione
morfologica, possibile anche soltanto nella stessa specie, non vuol dire che
esista necessariamente un’ibridazione effettiva tra 2 specie biologiche
diverse, o tra klepton diversi, anche se le probabilità che L & G abbiano
un’ibridabilità elevata mi appaiono molto alte, considerata l’elevata
variabilità dei loro morfi, dispersi invero per un ampio tratto di oceano
Atlantico West africano, il che favorirebbe l’insorgere di varietà locali.
Date le grandi dimensioni di L, dovrebbe
essere possibile l’ibridazione con quasi tutte le specie del Complex, tranne
forse le più piccole, tuttavia credo che M.
desjardini e pseudodesjardini non intervengano mai con L, mentre ritengo
assai possibili LXG, LXP, LXS, cioè che possano esserci ibridi, veri, oppure
solo morfologici apparenti, cioè in realtà vere specie e/o klepton, soltanto
nel quadrilatero delle specie L, G, P,
S, o meglio nel loro tetraedro morfologico, ponendo le caratteristiche tipiche
di ogni singola specie nei vertici di un immaginario tetraedro. S’intenda che L
assomiglia maggiormente a GXS, quindi si immagini il tetraedro con il vertice L
opportunamente spostato verso G & S. Vedere possibili ibridi collaterali
alla L tipica in alcune mie foto nella pagina Links.(GXS,
PXS, PXG).
In conclusione in alcuni casi di
morfologie miste rimane impossibile determinare l’identità precisa
dell’esemplare che si vuole classificare. Vedere ad esempio
l’Atlante italiano o l’Atlas inglese. Tuttavia in
questa imprecisione sta il bello della conchigliologia, ed anzi della
biodiversità individuale delle conchiglie.
Complex esteso a I = M. irrorata Menke, 1828 e cenno alle forme "albine" di G etc.
Per GXI probabilmente potrebbe
trattarsi solo di un'ibridazione morfologica senza ibridogenesi. Ma non si può
scartare del tutto la possibilità di esistenza di un improbabile klepton GXI. Mentre il klepton GXP
mi appariva molto probabile, diciamo al 90%, a motivo delle taglie e
similitudini conchigliari, per gli stessi motivi ritenevo dubitevole
l'ipotetico kl. GXI che, pur nella difficoltà del riconoscimento delle
caratteristiche morfologiche di G, interpretavo invece al 90% come una varietà
subspecifica solo di G: Marginella glabella
mutabilis Mattavelli 2005 (ed al 10% come vero Marginella klepton mutabilis),
inoltre nel 2005 non pensavo che potesse esistere la “specie” M. (kl. ?) visayae.
Nel 2021 la suddetta interpretazione è
stata sostituita, a percentuali ad intuito invertite, dal riconoscimento di Marginella (kl.?) visayae nuova “specie
sorella” di G & I, con diverse ipotesi biologiche, ancora da appurare
geneticamente.
Di “visayae” ovviamente non c’è traccia nel precedente elenco riepilogativo dei più grandi morfi più simili per ibridazioni morfologiche del Complex in analisi, per non complicare un elenco fatto in precedenza e già abbastanza complicato.
Indubbiamente la presenza di M. irrorata in Senegal e soprattutto Mauritania non deve essere affatto dimenticata, per le implicazioni con la presenza di “visayae”.
Sempre per le dimensioni avevo ritenuto quasi impossibile l'ibridazione PXI e IXL, tuttavia non è da trascurare l'esistenza della varietà Marginella irrorata pseudoirrorata, soltanto morfologicamente IXL, varietà analizzata in altra pagina del presente sito.
“pseudoirrorata” potrebbe infatti assumere qualche ruolo nell’esistenza di “visayae”, come analizzato nelle ipotesi biologiche dell’articolo Marginella (kl.?) visayae nuova “specie sorella”.
Un discorso integrativo meriterebbero
poi tutte le forme "albine", o meglio leucistiche, soprattutto di G, liquidate spesso in
letteratura come semplici varietà di G.
Per tali forme, oltre alla possibilità di introdurre anche l'ipotesi klepton forse GXI in forma leucistica (cioè M. kl. mutabilis albida, o meglio M. kl. visayae albida), penserei pure in alternativa a possibili indipendenti nuove specie, come ad esempio M. albida e M. tizianica, analizzate nella quadrilogia di articoli citati nella Nota novità del dicembre 2021, vedere links a capopagina, in particolare l’articolo Albinismo delle simil-glabella.
|
GXI = M. glabella"mutabilis" Mattavelli 2005 + kl.? Mattavelli 2011 (provvisoriamente) 33,7 x 19,2 mm - Malachia Forum Per "mutabilis" come var. di G (Mattavelli 2005) vedere marginella_glabella_mutabilis.htm |
Marginella glabella COMPLEX esteso a Marginella
irrorata
Per le innumerevoli varietà di G ed I rimando all'articolo web_4.htm, dove tuttavia non si supponeva l'ipotesi klepton.
Dalle foto
esposte si può pensare l'ipotesi klepton anche nell'ibridazione G X I, tuttavia richiamo particolare cautela su ogni concetto di
ibridogenesi. Vedere a fondo tabella M. visayae. |
GXP = M. glabella "antinea" Mattavelli 2005 + kl.? Mattavelli 2011 (provvisoriamente) 40,8 x 23,5 mm - Malachia Forum Per "antinea" come varietà di G (Mattavelli 2005) vedere problematica.htm |
|
I = Marginella irrorata Menke 1828 © Eddie Hardy |
G = Marginella glabella (Linneo 1758) Wikipedia Commons |
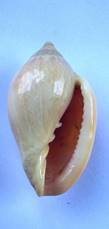
P = Marginella pseudosebastiani Mattavelli 2001 47,3 mm - sintipo Mattavelli |
|
S = Marginella sebastiani Marche Marchad & Rosso 1979 Wikipedia Commons |
D = Marginella desjardini Marche Marchad 1957 Wikipedia Commons |
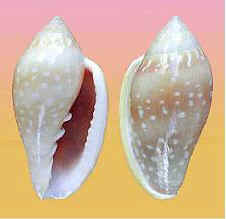
R = DXS = M. "pseudodesjardini" sp. Le Béon 2012 o kl. Mattavelli 2011? 49,1 mm - Roger Le Béon |
|
|
L = Marginella (kl.?) pseudoglabella - Nuovo sguardo ai mondi
di G & L - Descrizione
comparativa 2018 |
Considerare che “visayae” è stata
descritta nel 2021, sulla rivista MMM n° 113, in 2 forme, a puntinatura fine
(olotipo 41 mm, foto sotto a sinistra), oppure grossolana (41,4 mm, sotto a
destra, foto ©
Conchology Inc.). |
|
|
V = Marginella (Kl.?) Visayae Mattavelli, 2018- 2021 Descrizione negli
articoli: -
Prima supposizione di esistenza La forma di
sinistra in parte sostituisce M.
glabella mutabilis, mentre quella di destra in parte sostituisce M. glabelle antinea, rappresentate ad inizio
tabella. |
|










Tenere presente che la relaise iniziale di quest’articolo non considerava
né “pseudoglabella”, né “visayae”. Nel testo, derivato da
successivi aggiornamenti, potreste trovare qualche passata discrepanza con i
miei intendimenti più recenti, eventualmente scrivetemi.
Buona navigazione
Flavio Mattavelli
Per accedere a tutte le altre pagine del mio sito.